Hvordan oppstår celle-kompleksitet og kroppsplan
 Påstand: I biokjemi er det bare snakk om kjemiske reaksjoner.
Påstand: I biokjemi er det bare snakk om kjemiske reaksjoner.
Svar: Bruce Alberts, "Cellen som en samling av proteinmaskiner: Forbereder neste generasjon av molekylære biologer," Cell, 92 (6. februar 1998): 291-294) "Vi har alltid undervurdert celler. Utvilsomt gjør vi det fortsatt i dag. Men i det minste er vi ikke lenger så naive som vi var da jeg var utdannet student på 1960-tallet. Da, så de fleste av oss på celler som om de inneholdt et gigantisk sett av annenordens reaksjoner: molekyl A og B ble antatt å diffundere fritt, tilfeldig kollidere med hverandre for å produsere molekyl AB - og på samme måte for de mange andre molekylene som samhandler med hverandre. inne i en celle...
Men i stedet for å ha en celle dominert av tilfeldig kolliderende individuelle proteinmolekyler, vet vi nå at nesten alle større prosesser i en celle utføres ved samlinger av 10 eller flere proteinmolekyler. Og når den utfører sine biologiske funksjoner, samhandler hver av disse proteinsammensetningene med flere andre store proteinkomplekser. Hele cellen kan faktisk sees på som en fabrikk som inneholder et omfattende nettverk av sammenlåsende samlebånd, som hver består av et sett med store proteinmaskiner. "
Lenke.
Bilde 1. 'Proteinfabrikk'
Sjefredaktør for vitenskap (2009-2013). Dr Alberts arbeidet to seks års-perioder som president for National Academy of Sciences
Å forklare organismeform avhenger av å forklare hvordan organer, vev og celler dannes og danner form. På det laveste nivået av hierarkiet, avhenger det av dannelse av celler i en flercellet organisme av 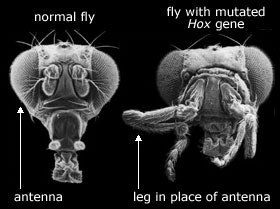 spesifikasjonen av:
spesifikasjonen av:
1. Morfogenese av forskjellige eukaryote celler, strukturer og former -herfra.
2. Bestemmelse og differensiering av celleskjebne (fenotype, eller hvilken celletype hver enkelt blir) -herfra.
3. Cellevekst og størrelse -herfra.
4. Utvikling og celledelings-antall: celler må programmeres til å stoppe selvreplikering etter riktig antall celledelinger -herfra.
5. Mekanismer for mønsterdannelse -herfra.
6. Hox-gener -herfra.
Bilde 2. Mutasjon i hox-gener fører til deformitet
7. Posisjon og plassering i kroppen. Dette er avgjørende. Lemmer som ben, finner, øyne osv. Må plasseres på rett sted -herfra.
8. Hvilken kommunikasjon det kreves for å kommunisere med andre celler, og oppsettet av kommunikasjonskanalene -herfra.
9. Sensoriske og stimuli funksjoner av celler -herfra.
10. Hvilke spesifikke nye regulatoriske funksjoner må celler tilegne seg
11. Når vil utviklingsprogrammet til organismen uttrykke genene til å gro ut de nye cellene under utvikling?
12. Endringsregulering i sammensetningen av cellemembranen og/eller utskilte produkter.
13. Spesifikasjon av cellecelleadhesjons-proteinene og hvilke som vil bli brukt i hver celle for å feste seg til nabocellene (det er 4 klasser)
14. Apoptose: programmering av tidsperioden cellen holder seg i live i kroppen, og når er det på tide å selvdestruere og erstattes av nyproduserte celler av samme slag
15. Sett opp hver celles spesifikke ernæringsbehov
16. Endring i celleform
17. Celleproliferasjon som er prosessen som resulterer i en økning i antall celler, og er definert av balansen mellom celledeling og celletap gjennom celledød eller differensiering.
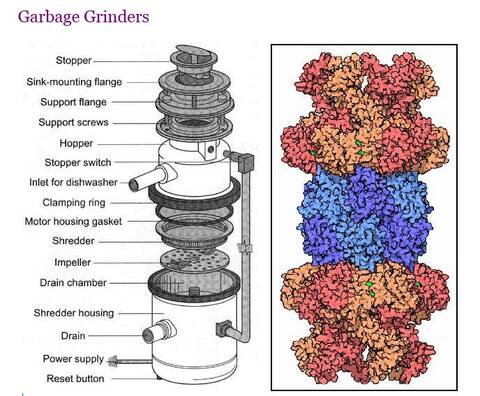
Bilde 3. Proteaosome -sml. avfallskvern
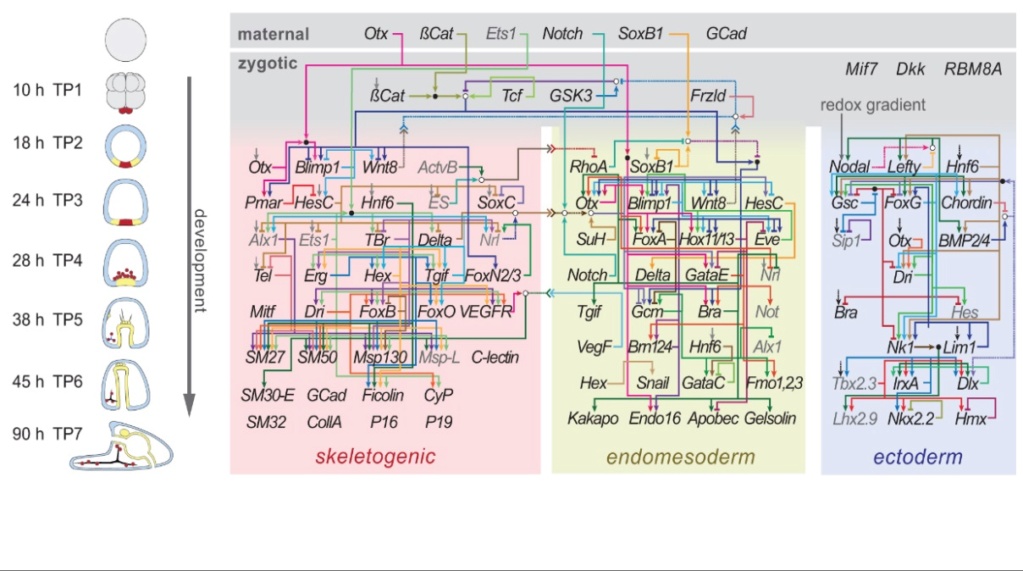
Hver celle i et embryo mottar molekylære signaler fra naboceller i form av proteiner, RNA og til og med overflate-interaksjoner. Nesten alle dyr gjennomgår en lignende sekvens av hendelser under veldig tidlig utvikling, en konservert prosess kjent som embryogenese. Under embryogenese eksisterer celler i tre kimlag og gjennomgår gastrulering. Et grunnleggende sett av de samme proteinene og mRNAene er involvert i embryogenesen.
Hvordan oppstod komplekse flercellede organismer, og hvordan oversettes genetisk og epigenetisk informasjon til en romlig organisert kroppsplan? For å tillate bygging av komplekse organer med intrikate mønstre for cellulær spesialisering, tas slike avgjørelser på et overcellulært nivå på en ikke-autonom måte-celle. Organismer krever ikke bare utviklingsmekanismer, men også strategier for å ordne og differensiere cellene, for å overleve og reprodusere.
En kropp er mer enn en samling av tilfeldig distribuerte celletyper. Utvikling innebærer ikke bare differensiering av celler, men også deres organisering i flercellede ordninger som vev og organer. Når vi observerer den detaljerte anatomien til et vev som den nevrale netthinnen i øyet, ser vi et intrikat og presist arrangement av mange typer celler. Hvordan kan materie organisere seg selv, slik at det blir en kompleks struktur som en lem eller et øye?
Bilde 4. Embryoutvikling -nettverk; -helt i starten
Det er fem store spørsmål til embryologer som studerer morfogenese:
 1. Hvordan dannes vev fra populasjoner av celler? For eksempel, hvordan henger nevrale netthinneceller seg fast til andre nevrale netthinneceller og blir ikke integrert i den pigmenterte netthinnen eller iriscellene ved siden av dem? Hvordan er de forskjellige celletyper i netthinnen (de tre forskjellige lagene med fotoreseptorer, bipolare nevroner og ganglionceller) ordnet slik at netthinnen er funksjonell?
1. Hvordan dannes vev fra populasjoner av celler? For eksempel, hvordan henger nevrale netthinneceller seg fast til andre nevrale netthinneceller og blir ikke integrert i den pigmenterte netthinnen eller iriscellene ved siden av dem? Hvordan er de forskjellige celletyper i netthinnen (de tre forskjellige lagene med fotoreseptorer, bipolare nevroner og ganglionceller) ordnet slik at netthinnen er funksjonell?
2. Hvordan er organer konstruert av vev? Øyets netthinne dannes i en nøyaktig avstand bak hornhinnen og linsen. Netthinnen ville være ubrukelig hvis den utviklet seg bak et bein eller midt i nyrene. Videre må nevroner fra netthinnen komme inn i hjernen for å stimulere regionene i hjernebarken som analyserer visuell informasjon. Alle disse tilkoblingene må være presist ordnet.
3. Hvordan dannes organer på bestemte steder, og hvordan når migrerende celler sine destinasjoner? Øynene utvikler seg bare i hodet og ingen andre steder. Hva hindrer et øye i å dannes i et annet område av kroppen? Noen celler, for eksempel forløperne til pigmentcellene, kimcellene og blodcellene våre, må reise lange avstander for å nå sine endelige destinasjoner. Hvordan blir celler bedt om å reise langs bestemte ruter i våre embryonale kropper, og hvordan blir de bedt om å stoppe når de har nådd de riktige destinasjonene?
4. Hvordan vokser organer og deres celler, og hvordan koordineres deres vekst gjennom hele utviklingen? Cellene i alt vev i øyet må vokse på en koordinert måte hvis man skal se. Noen celler, inkludert de fleste nevroner, deler seg ikke etter fødselen. Derimot tapper tarmene stadig celler, og nye tarmceller regenereres hver dag. Den mitotiske frekvensen til dette vevet må reguleres nøye. Hvis tarmen genererte flere celler enn den slo av, kan den produsere tumor-utvekster. Hvis den produserte færre celler enn det slo av, ville den snart bli ikke-funksjonell. Hva styrer frekvensen av mitose i tarmen?
5. Hvordan oppnår organer polaritet? Hvis man skulle se på et tverrsnitt av fingrene, ville man se en viss organisert samling av vevsben, brusk, muskler, fett, lærhud, overhud, blod og nevroner. Ser man på et tverrsnitt av underarmen, vil man finne den samme samlingen av vev. Men de er ordnet veldig forskjellig i forskjellige deler av armen. Hvordan kan det være at de samme celletyper kan ordnes på forskjellige måter i forskjellige deler av samme struktur? Alle disse spørsmålene gjelder aspekter ved celleoppførsel. Det er to hovedtyper av cellearrangementer i embryoet: epitelceller, som er tett forbundet med hverandre i plater eller rør, og mesenkymale celler, som ikke er koblet til hverandre og som fungerer som uavhengige enheter. Morfogenese oppstår gjennom et begrenset repertoar av variasjoner i cellulære prosesser innenfor disse to typer ordninger:
Forskjeller i regulatorisk DNA forklarer morfologiske forskjeller
 Dyr inneholder de samme essensielle celletyper, har en lignende samling av gener og deler mange av de molekylære mekanismene for mønsterdannelse. Men hvordan kan vi sammenholde dette med de radikale forskjellene vi ser i kroppsstrukturene til dyr som er så forskjellige som en orm, en flue, en frosk og en mus? På en generell måte ser disse forskjellene vanligvis ut til å gjenspeile forskjeller i det regulatoriske DNAet som spiller inn komponentene i det konserverte grunnleggende settet med deler. Vi må nå undersøke bevisene litt nærmere. Når vi sammenligner dyrearter med lignende grunnleggende kroppsplaner - for eksempel forskjellige virveldyr, for eksempel fisk, fugler og pattedyr, finner vi at tilsvarende gener vanligvis har lignende sett med regulatoriske elementer: De regulatoriske DNA-sekvensene har blitt bevart og er gjenkjennelig homolog i de forskjellige dyrene. Det samme gjelder hvis vi sammenligner forskjellige arter av nematodeormer eller insekter. Men når vi sammenligner virvelløse reguleringsregioner med ormer eller fluer, er det vanskelig å se noen slik likhet. De proteinkodende sekvensene er umiskjennelig like, men de tilsvarende regulatoriske DNA-sekvensene virker stort sett veldig forskjellige, noe som tyder på at forskjellene i kroppsplaner hovedsakelig gjenspeiler forskjeller i regulatorisk DNA.
Dyr inneholder de samme essensielle celletyper, har en lignende samling av gener og deler mange av de molekylære mekanismene for mønsterdannelse. Men hvordan kan vi sammenholde dette med de radikale forskjellene vi ser i kroppsstrukturene til dyr som er så forskjellige som en orm, en flue, en frosk og en mus? På en generell måte ser disse forskjellene vanligvis ut til å gjenspeile forskjeller i det regulatoriske DNAet som spiller inn komponentene i det konserverte grunnleggende settet med deler. Vi må nå undersøke bevisene litt nærmere. Når vi sammenligner dyrearter med lignende grunnleggende kroppsplaner - for eksempel forskjellige virveldyr, for eksempel fisk, fugler og pattedyr, finner vi at tilsvarende gener vanligvis har lignende sett med regulatoriske elementer: De regulatoriske DNA-sekvensene har blitt bevart og er gjenkjennelig homolog i de forskjellige dyrene. Det samme gjelder hvis vi sammenligner forskjellige arter av nematodeormer eller insekter. Men når vi sammenligner virvelløse reguleringsregioner med ormer eller fluer, er det vanskelig å se noen slik likhet. De proteinkodende sekvensene er umiskjennelig like, men de tilsvarende regulatoriske DNA-sekvensene virker stort sett veldig forskjellige, noe som tyder på at forskjellene i kroppsplaner hovedsakelig gjenspeiler forskjeller i regulatorisk DNA.
Bilde 5. Ulike celletyper
Selv om variasjoner i proteinene i seg selv også bidrar, vil forskjeller i regulatorisk DNA være nok til å generere radikalt forskjellige vev og kroppsstrukturer selv om proteinene var de samme. Det er ennå ikke mulig å spore de genetiske trinnene som har ført til alt det spektakulære mangfoldet av dyr. Studier av både nært beslektede dyrepopulasjoner og plantepopulasjoner hvis medlemmer har forskjellige morfologier, har avdekket at dramatiske utviklingseffekter kan skyldes subtile endringer i regulatorisk DNA. Et godt studert eksempel er et morfologisk mangfold som finnes i stingsild. Etter at den siste istiden endte for rundt 10 000 år siden, koloniserte marine stingsild mange nydannede ferskvannselver og innsjøer. Marine stingsild har skarpe pigger ut fra bekkenskjelettet. Disse piggene antas å bidra til å beskytte fisken mot rovdyr med myk munn. I motsetning til dette har flere populasjoner med ferskvanns stingsild mistet disse piggene, vanligvis i innsjøer som mangler slike rovdyr. De forskjellige morfologiene gjenspeiler forskjeller i kontroll over uttrykket av en transkripsjonsregulator kalt Pitx1. Mens marine stingsild uttrykker Pitx1-genet i bekkenbenforløpercellene som vil danne piggene, har ferskvanns stingsild mistet dette uttrykket som et resultat av en endring på Pitx1-locus. Disse endringene ligger ikke i kodingssekvensen. I stedet er hver en liten sletting av en blokk med tilstøtende regulatorisk DNA som styrer Pitx1-ekspresjon spesifikt i bekkencellene.
Morfologisk mangfold i stingsild er forårsaket av endringer i reguleringselementer.
(A – D) Bekken pigger er tilstede i marine (A) men ikke i ferskvanns (C) populasjoner. Tilsvarende uttrykkes Pitx1 i bekkenområdet i marine (B), men ikke i ferskvannsfisk (D). Mangelen på uttrykk i bekkenområdet hos ferskvannspopulasjoner er forårsaket av mutasjoner i et forsterkningselement. Andre forsterkere og uttrykkssteder for Pitx1 er de samme i marin og ferskvanns stingsild.
 Bilde 6:Stingsild -marin (øverst) -ferskvanns stingsild (nederst)
Bilde 6:Stingsild -marin (øverst) -ferskvanns stingsild (nederst)
Pitx1-proteinet har viktige funksjoner andre steder i kroppen, slik at DNA-sekvensene som koder for dette proteinet, må beholdes. Det regulatoriske DNA som er ansvarlig for Pitx1-uttrykk på disse andre stedene, er også uendret i de to populasjonene av stingsild.
Morfogenese av eukaryote celler, struktur og form
Herfra.
Å svare på spørsmålene om hvordan celler, vev og organismer dannes, går foran spørsmålet om hvordan de til slutt kan mangfoldiggjøres, utvikle seg, forandre seg og forvandle seg fra en art til en annen gjennom en makroevolusjonær primær spesiasjon overgangssone, der nye organismeegenskaper oppstår, som vinger, øyne, ører, ben, armer og så videre. Faktum er at vitenskapen fremdeles er langt fra å kunne svare på det spørsmålet på en uttømmende måte. Minst 23 epigenetiske koder og språk er vitenskapelig kjent. De bidrar alle til organismeutvikling på en avgjørende måte.
Et Google-søk vil demonstrere at det ikke er noen vitenskapelige artikler som viser løsning på bare en av disse kodene. Vitenskapen har ingen anelse om hvordan og hvor de lagres, og heller ikke betydningen av kodifisering, tilordning og regler, språk eller kryptering. Vitenskapen vet at de eksisterer gjennom eksperimentelle tester, men dypere detaljer er ikke kjent.
Se flere detaljer her.
Oversettelse og bilder ved Asbjørn E. Lund